Можливості застосування CRISPR/Cas9 для генної терапії вірусу імунодефіциту людини
Вірус імунодефіциту людини (ВІЛ) атакує клітини імунної системи, а саме CD4+ Т-хелпери, залишаючи організм людини на поталу збудникам найрізноманітніших інфекцій. Проте, наскільки б неймовірним не здавалося б вам таке твердження, на допомогу людині в найбільш скрутну годину приходить імунна система бактерії. Згаданий вірус оповитий туманною аурою різноманітних міфів: ніби-то його випустили ЦРУ для контролю кількості населення Африки, чи це зробили китайці для з тією самою метою проти США, а інтернет-повідомлення про винайдення чудодійних ліків проти ВІЛ з’являються чи не щогодини. У той же час, щороку в світі публікуються тисячі наукових робіт із дослідженнями наноскопічного вбивці. Але вірус із холоднокровною байдужістю ставиться до тієї шани, котру віддає йому вищий примат виду Homo sapiens sapiens, і тишком-нишком робить свою вбивчу справу. Ми ж з вами залишимо осторонь спекуляції та змови, які, звичайно, не мають нічого спільного із реальністю, і критично оцінимо можливості застосування бактеріальної системи захисту проти фагів CRISPR/Cas9 в боротьбі із ВІЛ/СНІДом, мимохідь смакуючи найкрасивіші моменти з історії генної терапії цієї недуги.
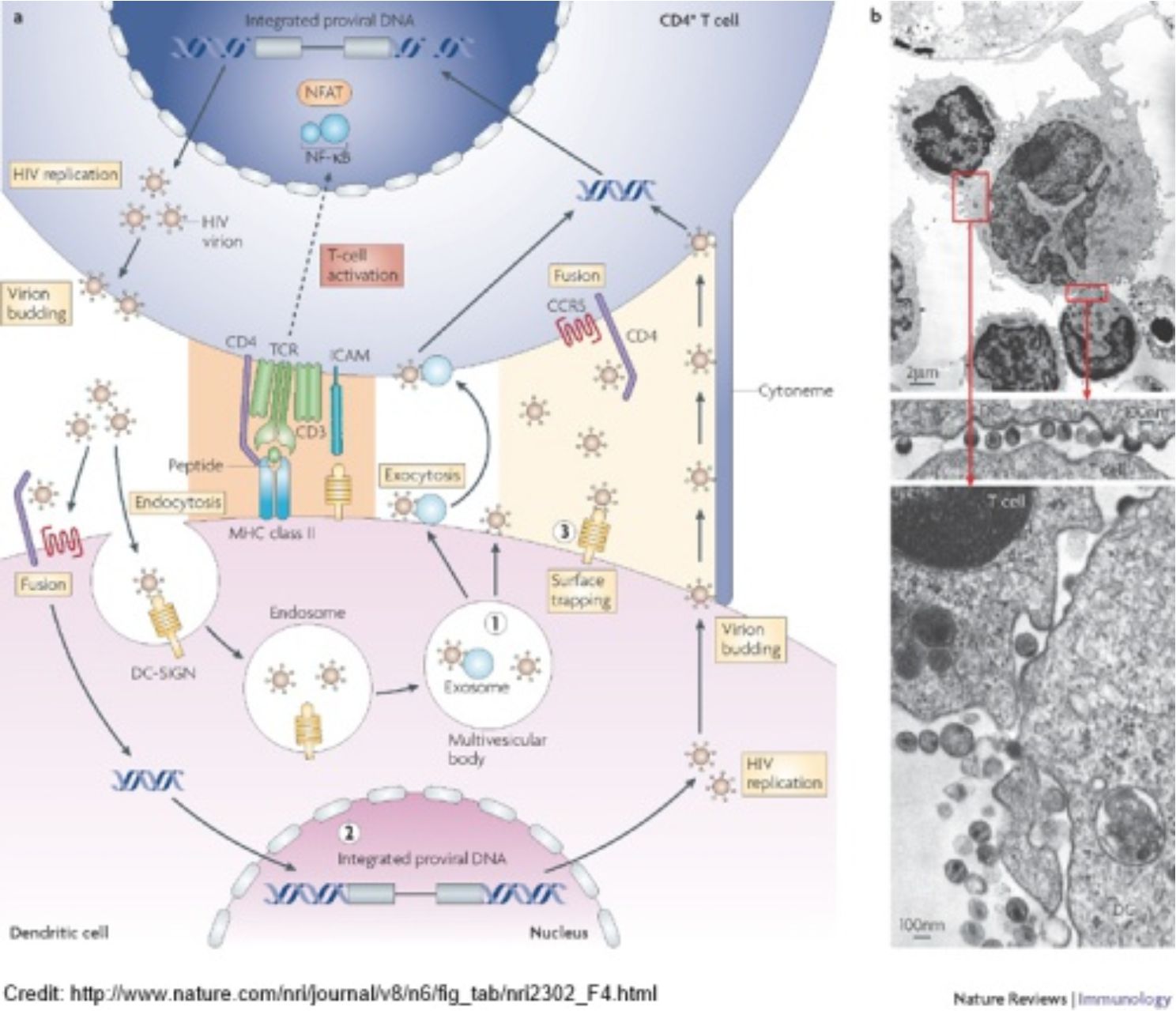
В середині 90-х років минулого сторіччя стало відомо, що ВІЛ використовує рецептор CD4 та корецепторCCR5 для входження в клітину. Цим, власне, зумовлена його специфічність до Т-хелперних клітин та деяких дендритних клітин. ВІЛ є ретровірусом, це означає, що він існує як в цитоплазмі клітини у вигляді РНК, так і в стані інтегрованому в геном клітини, тож інфекція протягом тривалого часу протікає латентно. Проте в якийсь момент віруси масово переходять від лізогенної до літичної фази розвитку, що має наслідком швидку загибель клітин-носіїв, власне, CD4+ Т-клітин та дендритних клітин. Оскільки останні виконують інтегральну роль в розвитку імунних реакцій людини, то виведення їх з ладу веде за собою розвиток імунодефіциту. На рисунку нижче підсумовано шляхи передачі ВІЛ від клітини до клітини.
CCR5 (Cysteine-cysteine chemokine receptor 5) є рецептором так званих хемокінів – молекул, які беруть участь в підтриманні запалення та спрямовуванні клітин до «гарячої точки імунного протистояння». Протягом останнього часу саме він привернув увагу вчених як можлива ціль генної терапії ВІЛ/СНІДу, і на це були свої причини. По-перше, CCR5 є абсолютно необхідним для входження ВІЛ всередину клітини, що підтверджується кількома дослідженнями, основні з яких приведені нижче. По-друге, його роль в регуляції імунітету, як видається, не є суттєвою – його функції дубльовані іншими рецепторами хемокінів. На відміну від більшості генів, втрата експресії CCR5 не викликає суттєвих порушень. Отже, можна без особливого ризику для організму вимкнути ген, що кодує протеїн CCR5, і таким чином позбавити вірус імунодефіциту можливості передаватися від клітини до клітини.
Приклад 1. Резистентність до вірусу носіїв гомозиготної мутації CCR5-Δ32, яка є делецією 32-х пар основ з 794 по 825 нуклеотид. Така делеція по суті є нонсенс мутацією, призводячи до зсуву рамки зчитування і появи передчасного стоп-коду. Відповідно, мРНК рецептору не транслюється, а піддається нонсенс-опосередкованому розпаду (nonsense-mediated decay). Тож, такі індивіди не експресують CCR5 на поверхні імунних клітин, почуваються добре і не хворіють на СНІД. За даними генетиків носіями такої мутації є до 18 % жителів Європи, а в її північній частині до 20 %. Існує припущення, що висока частота такої мутації спричинена селективним тиском на людську популяцію таких патогенів як Yersinia pestis(збудник чуми) та Variola virus (збудник віспи) протягом тривалого часу.
Приклад 2. У 2008 році пацієнту із ВІЛ/СНІД, в якого на фоні імунодефіциту спостерігався розвиток гострої лімфобластної лейкемії, було здійснено трансплантацію кісткового мозку від здорового донора, гомозиготного за CCR5-Δ32 мутацією. Операція пройшла успішно, і заразом із лейкемією його таким чином вилікували від ВІЛ. Точніше сказати, у нього спостерігався різкий спад кількості вірусу в крові, але, як відомо, пацієнти після трансплантації від неродинного донора повинні постійно приймати імуносупресанти для попередження можливого відторгнення. Тож, така цинічна іронія долі.
Приклад 3. Під час дослідження хворих на гемофілію, які отримували фактор VIII із сироватки крові людини, виявилося, що вони були несприйнятливі до ВІЛ, який також випадково містився в цій сироватці. Дослідження проводилося в середині 80-х, коли про ВІЛ лишень дізналися – обов’язкового скринінгу донорів сироватки тоді ще не проводили. З’ясувалося, що у пацієнтів-гемофіліків були підвищені рівні експресії бета-хемокінів. Останні конкурували із лентівірусом (у тому числі ВІЛ) за зв’язування з CCR5 рецептором, не даючи йому шансу пробратися в клітину.
Станом на 2007 рік Національний Інститут Здоров’я Сполучених Штатів сумарно профінансував три десятки лабораторій на суму 10 мільйонів докризових доларів, поставивши їм завдання розробити ефективну генну терапію від СНІДу. Це не враховуючи 14 мільйонів, які отримав нобелівський лауреатДевід Балтімор на розробку супер-антитіла проти вірусу імонудефіциту. Тож, як бачимо, американські вчені достатньо щільно взялися за цю проблему.
Загалом усі підходи генної терапії СНІДу мають кілька схожих етапів: вчені забирають клітин крові пацієнта з коровотоку або безпосередньо з кісткового мозку, збагачують препарат на клітини, на які саме націлений вірус, вносять генетичні модифікації в них різними шляхами, та повертають клітини назад пацієнту. Головні відмінності полягають в типі генного вектора, що використовується для здійснення модифікації, а також в способі культивування клітини перед процедурою трансфекції, що в свою чергу залежить від ступеню диференційованості клітин, які беруться – попередники мієло- та лімфопоезу, які вже стали на шлях диференціювання (так звані “committed progenitor cells“), або ж недиференційованістовбурові клітини кровотворення. Ключем до успішності терапії є як ефективна трансформація великої кількості клітин, так і виживаність клітин після повернення до організму пацієнта. Слід зазначити, що існують численні стратегії генної терапії ВІЛ, але, за неможливістю охопити їх усі, ми зосередимось саме на тих, що спрямовані проти гену CCR5.
Коротко про основні здобутки на цій ниві станом на почак 2000-х. Steinberger et al. (2000) розробив внутрішньоклітинне одноланцюгове антитіло (так зване intrabody), яке зв’язує протеїн CCR5 під час його транспорту по секреторній системі клітини, таким чином затримуючи його і не даючи досягнути поверхні клітини. В теорії це мало б викликати набуття клітиною CCR5–/– фенотипу навіть без втручання в структуру самого гену. Strizki et al. (2001) синтезували малу молекулу, що блокує CCR5-опосередковане входження вірусу в клітину. Це зовсім не генна терапія, але вона також спрямована проти згаданого хемокінового рецептору. Препарат продемонстрував 50-60 відсоткову біодоступність у разі орального застосування і час напіврозпаду в сироватці до шести годин на тваринних моделях. Також були повідомлення (Qin et al., 2003) про застосування малих шпилькових РНК (siRNA) проти мРНК CCR5. Був розроблений лентівірусний вектор для доставки послідовності siRNA до імунних клітин. За іронією долі використовується той самий вірус СНІДу тільки в зміненій формі.
Але усі перелічені стратегії не виявилися достатньо ефективними (в іншому разі, з огляду на роки публікацій, ми б вже давно знали про існування ліків від СНІДу). Фармакологічне подавлення рецептору потребує постійного прийому достатньо великих доз препарату, додатково до того «коктейлю», який зазвичай приймається хворим на імунодефіцит. До того ж у підсумку виникає резистентність вірусу. Підходи з intrabody та siRNA передбачають процедуру вставляння нових генів у геном клітин крові людини, що почасти пов’язано із двома серйозними проблемами – низькою ефективністю модифікації та ризиком для життя пацієнта. По-перше, принципово неможливо досягнути 100% трансформації усіх клітин (Т-хелперів та дендритних клітин). Насправді, вірогідність вставки гену навіть шляхом гомологічної рекомбінації настільки низька, що просто не дозволить отримати потрібну кількість модифікованих клітин, де intrabody або siRNA експресувалися б достатньо інтенсивно для успішного пригнічення CCR5. Але це, ще півбіди, оскільки навіть така непевна вставка може втрачатися (через ще одну рекомбінаційну подію або епігенетичний сайленсінг) протягом першого ж року після терапії. І в такому випадку синтез функціонального CCR5 рецептору буде поновлюватися. По-друге, виникає небезпека інсерційного мутагенезу. Наприклад, у 2005 році з’явилося повідомлення про розвиток лейкемії у дітей, що отримували генну терапію від СНІДу (як вектор використовувалися мишачі ретровіруси). Лентівірусні вектори працюють краще і вважаються безпечнішими за ретровіруси лейкемії мишей, проте у разі використання ВІЛ-подібних вірусів для доставки генетичних конструкцій виникає ще одна страшна небезпека: між хвороботворним та терапевтичним вірусами може відбутися рекомбінація, наслідки якої складно передбачити. На рисунку нижче, узагальнено усі наявні наразі стратегії пригнічення експресії рецептору CCR5.
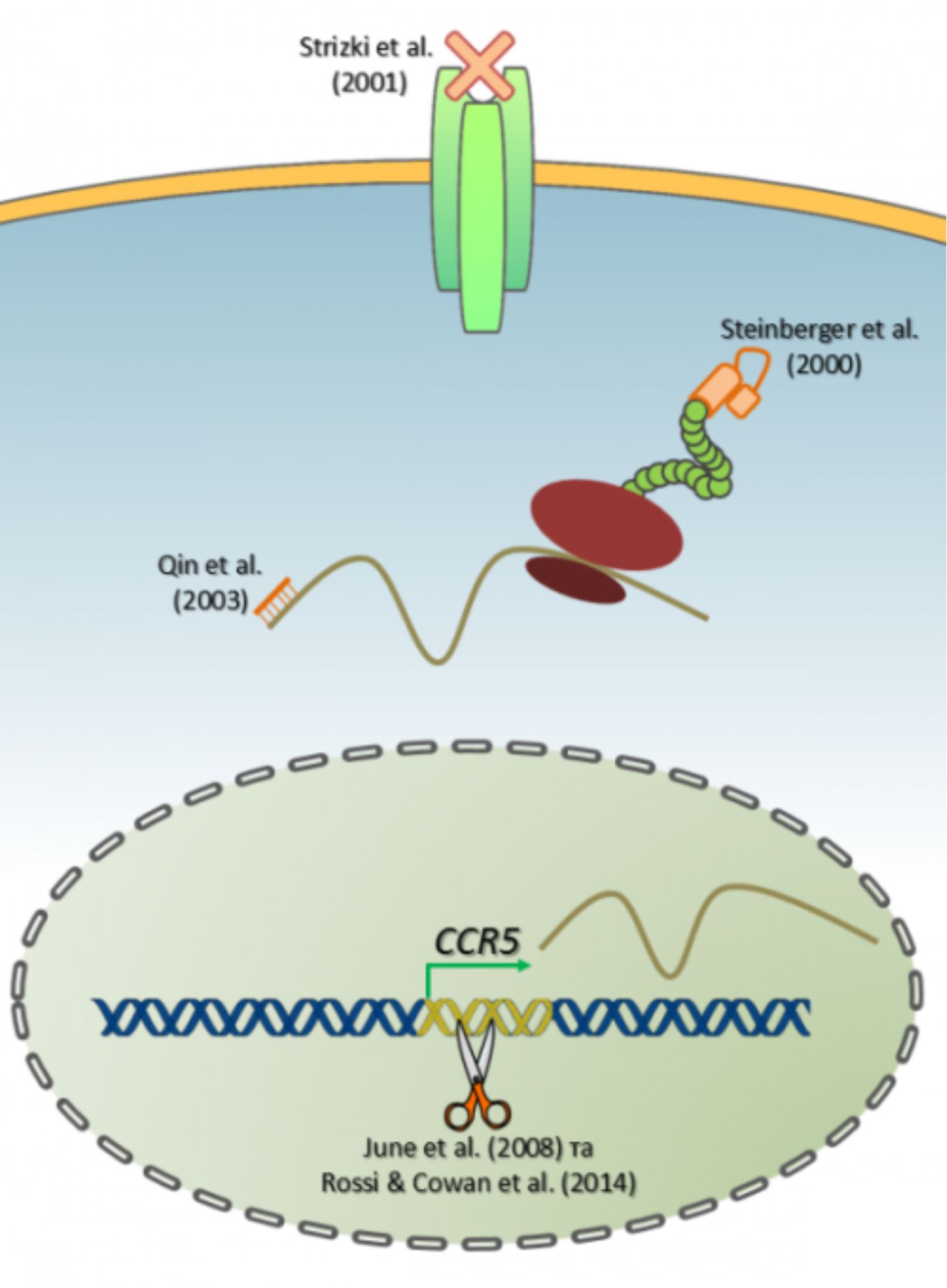
У 2005 на обрії з’явилася технологія, покликана подолати усі вищезазначені проблеми, її назва –геномне редагування. Винайдення модульних едонуклеаз із програмованою специфічністю, таких як нуклеази із ДНК-зв’язуючими доменами «цинкові пальці» (zinc-finger nucleases, ZFN) та трансактиватор-подібні ефекторні нуклеази (Transcription activator-like effector nucleases, TALENs), надало можливість напряму маніпулювати із генами безпосередньо на клітинній ДНК. За принципом «зламати легше, ніж збудувати», такий підхід значно спрощує нокаутування генів – достатньо лише нетривалої експресії ZFN в клітині для того, щоб незворотньо пошкодити певний клітинний ген. Тож, не дивно, що цією стратегією вирішили скористатися борці зі СНІДом. Взагалі, редагування геному, ZFN та TALENs достойні окремого огляду на сторінках Моєї Науки {може іншим разом ;)}.
Перша спроба геномного редагування за допомогою ZFN нуклеаз у Т-клітинах була здійснена June et al. (2008). Тимчасова експресія в складі аденовірусного вектора ZFN протеїну, націленого на ген CCR5, викликала ефективне і стабільне нокаутування цього гену, що спадкувалося в ході подальшої проліферації цих клітин. Цікавою знахідкою було те, що нокаут CCR5 надає селективну перевагу CD4+ T-клітинам у разі їх культивування із вірусом імунодефіциту in vitro. Надалі вчені спостерігали схожі результати in vivo, використовуючи мишачу модель ВІЛ. Вони продемонстрували, що пересадження ВІЛ-інфікованим мишам ZFN-модифікованих CD4+ T-клітин призводить до значного зменшення вірусного навантаження на фоні зростання кількості CCR5–/– Т-клітин в довгостроковій перспективі. Не гаючи часу, дослідники взялися до реалізації цієї стратегії в клініці. В період з травня 2009 по липень 2012 було залучено 12 ВІЛ/СНІД-пацієнтів, а результати дослідження були опубліковані нещодавно (березень 2014) в журналі New England Journal of Medicine. Подібно до попереднього експерименту, спочатку в пацієнтів було зібрано CD4+ T-лімфоцити, потім в них було виконано нокаут CCR5 гену шляхом тимчасової експресії специфічної ZFN нуклеази із подальшим поверненням модифікованих клітин назад до кровотоку пацієнтів. За 4 тижні після вдалого перенесення клітин 6 з 12-ти пацієнтів були зняті із супроводжуючої антиретровірусної терапії з метою дослідження динаміки CD4+ Т-хелперних клітин. Виявилося, що за 12 тижнів без супутньої терапії кількість Т-хелперів дикого типу різко скоротилася на фоні незначущих коливань кількості модифікованих Т-хелперів, що знову свідчило про позитивну селекцію щодо CCR5–/–. Протягом 254 днів випробувань показники вірусного навантаження в крові пацієнтів впали на три порядки (в 1000 разів), але повного очищення від вірусу імунодефіциту вдалося досягти лише у одного пацієнта. Як виявилося пізніше, він був гетерозиготним за CCR5-Δ32 мутацією, тож ефективність подвійного нокауту (CCR5–/–) в Т-клітинах цього індивіда була вдвічі вищою, ніж у інших. Це зайвий раз підтверджує важливість згаданого корецептора як специфічної цілі для генної терапії ВІЛ/СНІДу.
У світлі таких захопливих наукових результатів, міжнародна дослідницька група (з 12-ти американських та одного шведського інститутів) вирішила піти далі і застосувати іншу ефективнішу систему геномного редагування для видалення CCR5 гену в гемопоетичних стовбурових клітинах людини. Ось тут на передній план виходить анонсована раніше імунна система бактерії. Її застосуванню в якості інструмента генетичних маніпуляцій передував тривалий період (30 років) вивчення її природи. Спершу увагу вчених привернули чудернацькі послідовності в геномі багатьох бактерій – так звані групи коротких паліндромних повторів, перемежованих із унікальними послідовностями (Clustered Regularly Interspaced Short Palindromic Repeats, CRISPR). У ході досліджень з’ясувалося, що вони є нічим іншим, як записаною до бактеріального геному пам’яттю про бактеріофагів, які колись потрапляли в клітину. У разі повторної атаки вірус-специфічні некодуючі РНК (так звані crRNA), що транскрибуються з CRISPR-локусу, допомагають спеціальним ендонуклеазам знаходити на знищувати фагову ДНК шляхом утворення дволанцюгових розривів, реалізуючи таким чином набутий імунітет. Механізм трохи подібний до РНК інтерференції, але той факт, що він спрямований саме проти дволанцюгової ДНК, а не РНК, робить CRISPR систему безцінним інструментом генно-інженерних досліджень, а особливо стала у пригоді науковцям CRISPR/Cas9 Streptococcus pyogenes. Тільки за 2013 рік з’явилося щонайменше з десяток публікації з її використанням у трійці найвпливовіших біологічних журналів (Nature/Cell/Science). Оскільки, немає змоги вставити сюди детальний опис CRISPR/Cas9, запрошую читачів докладніше ознайомитися (рідною мовою!) з історією дослідження, молекулярним механізмом дії та типами маніпуляції із геномом тут.
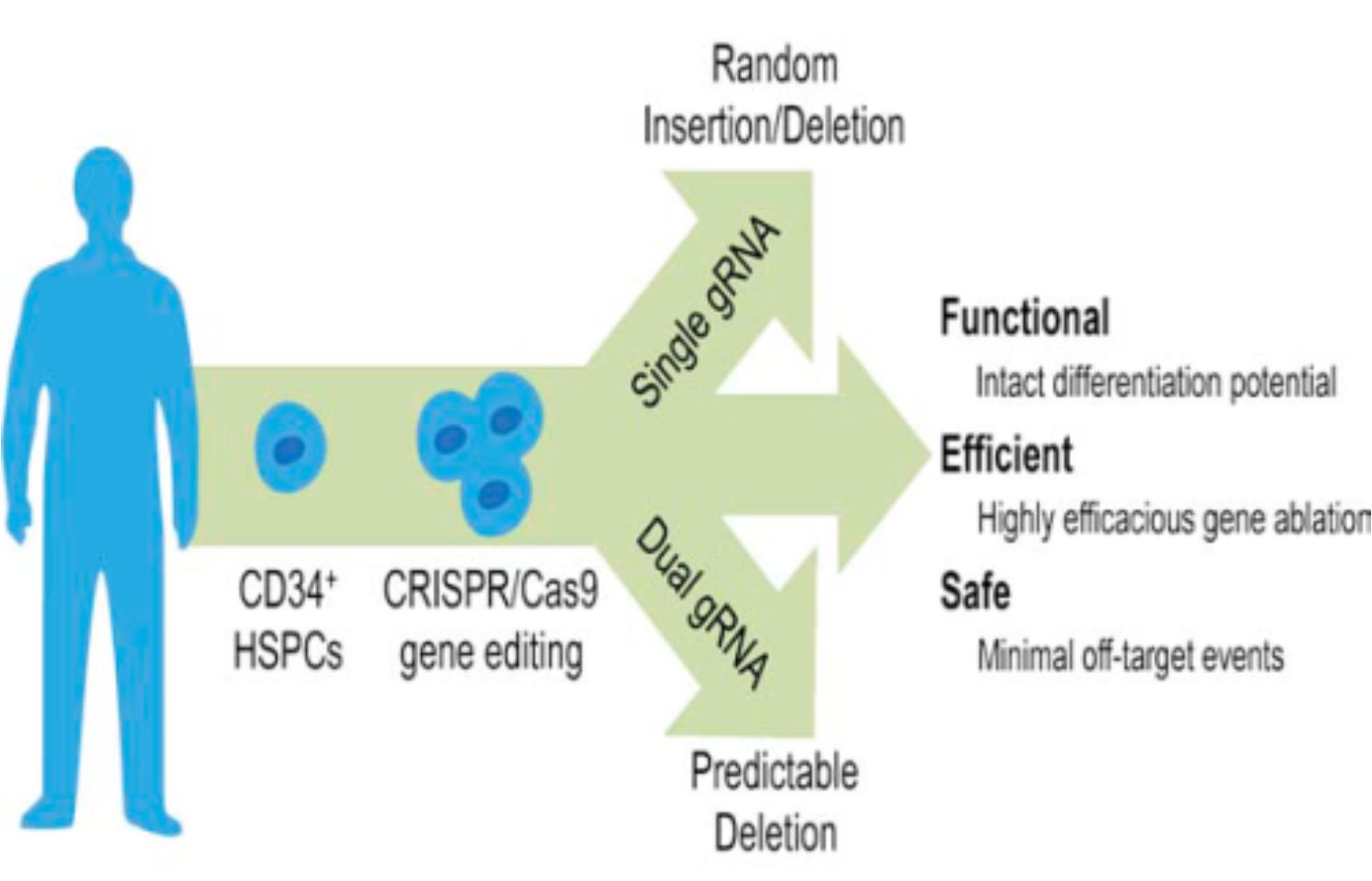
Отже, судячи зі статті, опублікованій в Cell Stem Cell, вченим вдалося довести, що CRISPR/Cas9 в комбінації із CCR5-специфічними crRNA є ефективною в здійсненні нокауту гену CCR5 в недиференційованих клітинах кровотворення, так званих гемопоетичних стовбурових клітинах (ГСК) людини. Відповідно до схеми експерименту, було виконано нуклеофекцію ГСК двома плазмідами, одна з яких несла послідовність ендонуклеази Cas9 під сильним цитоменаловірусним промотором, а друга – послідовність однієї або двох crRNA (під U6 промотором РНК-полімерази ІІІ), що виконували наведення ендонуклеази на ген CCR5. Важливо відзначити, що такий підхід ґрунтувався на тимчасовій експресії зазначених генетичних конструкцій подібно до ZFN-стратегії, але не передбачав використання вірусів для переносу генів, що значно підвищує безпечність такої процедури у разі її застосування із терапевтичною метою. В ході аналізу даних було встановлено, що найбільша ефективність нокауту спостерігалася у разі одночасного використання двох crRNA, спрямованих проти різних частин генуCCR5, що мало наслідком делецію або інверсію фрагмента, який містився між сайтами розрізання, і в підсумку призводило до нокауту. Принципово важливою знахідкою було те, що після модифікації ГСК зберігали здатність диференціюватися у всі типи клітин крові: in vitro утворювали різні типи колоній на метилцелюлозі, а in vivo спостерігалася успішна репопуляція кісткового мозку та селезінки NOD/SCIDмишей. Разом із тим CCR5–/– фенотип передавався їхнім нащадкам – спеціалізованим клітинам крові. Також глибоке секвенування отриманих клітин виявило низький рівень неспецифічності (або так званих позацільових ефектів – off-targeteffects), але в одному випадку таки було встановлено неспецифіку: під «гарячу руку» Cas9 потрапив ген CCR2, що мав гомологічні ділянки із цільовим геном. Проте, як запевняють вчені, якщо дуже ретельно підходити до дизайну crRNA, такі випадки можна легко попередити. Тож, побачимо, чи отримає дане дослідження продовження у вигляді клінічних випробувань. Теоретично, у разі модифікації стовбурових клітин крові нокаутований алель стабільно відтворюється у всіх клітинах крові протягом більш тривалого часу в порівнянні із модифікацією зрілих клітин крові. У попередньому дослідженні із ZFN-стратегією пацієнти спостерігалися лише протягом 300 днів, а що відбувалося із ними потім – історія, як то кажуть, замовчує. Ефективний нокаут в стовбурових клітинах по ідеї дозволить забезпечити пацієнта нечутливою до вірусу СНІДу імунною системою аж до старості. Нижче подано графічний огляд з оригінальної публікації в Cell Stem Cell.
Для того, щоб трохи збалансувати оптимізм – коротко про можливі недоліки та перестороги такого підходу. По-перше, існує певна лабільність env-протеїну вірусу імунодефіциту, тож існує ймовірність, що внаслідок селекції вірус почне використовувати інші схожі хемокінові рецептори для входження в клітину. Зокрема є відомості, що ВІЛ не цурається CXCR4, і хтозна які ще в нього є можливості варіабельності? По-друге, слід поглянути на рисунок, поданий на початку, який зображує шляхи передачі ВІЛ, щоб зрозуміти, що CD4/CCR5 — це основний, проте не єдиний спосіб входження вірусу до імунних клітин. В умовах селективного тиску допоміжні шляхи певно стануть основними. Тож, вірогідно, такий підхід до терапії ВІЛ/СНІДУ не буде панацеєю, а натомість дозволить виявити нові досі невідомі аспекти біології вірусу. По-третє, досі вичерпно не досліджені всі можливі функції рецептору CCR5. Те, що його нокаут суттєво не впливає на обмежену когорту пацієнтів, ще не означає безпроблемну застосовуваність такої терапії до всіх без винятку хворих. Знову-таки: питання до клініцистів, які володіють якісними і кількісними методами оцінки й передбачення ефективності певних методів лікування. А нам – біологам – залишається милуватися красою та складністю природи, і застосовувати увесь арсенал своєї уяви і винахідливості для пошуку відповідей на питання, що вона перед нами ставить.
Даниїл Грибчук (molecancereviw) для Моєї Науки
Запрошую продовжити читання про можливості використання CRISPR/Cas9 для швидкого моделювання канцерогенних мутацій in vivoна сторінках мого блогу:
Шляхами непокірних клітин | Молекулярно про Рак >>
Посилання
Online Mendelian Inheritance in Man®, entry #601373 (chemokine, cc motif, receptor 5; ccr5)
Cohen J. Building an HIV-Proof Immune System. Science. Vol. 317, No. 5838 (August, 2007): 612-614.
Holmes M.C., JuneC.H.,et al. Establishment of HIV-1 resistance in CD4+ T cells by genome editing using zinc-finger nucleases. Nature Biotechnology. Vol. 26, No. 7 (July, 2008): 808-816.
JuneC.H., Bruce B.L., et al. Gene Editing of CCR5 in Autologous CD4 T Cells of Persons Infected with HIV. The New England Journal of Medicine. Vol. 370, No. 10 (March, 2014): 901-910.
Rossi D.J., Cowan C.A., et al. Efficient Ablation of Genes in Human Hematopoietic Stem and Effector Cells using CRISPR/Cas9. Cell Stem Cell. Vol. 15 (November, 2014): 643–652.