 Віруси є визначними рушіями глобальної біогеохімії та етіологічними агентами багатьох хвороб. Вони також є переможцями у грі життя: на планеті вірусів більше, ніж клітинних організмів, і вони представляють більшу частину генетичного розмаїття планети. У дійсності, є підстави вважати життя вірусним інкубатором. Однак, більшість екологічних та еволюційних теорій, що були розвинуті й продовжують розвиватися, не беруть до уваги віросферу. Це означає, що ці теорії потребують повторної інтерпретації у світлі знань про віруси, або ж ми маємо розробити нову теорію з вірусологічної точки зору. У цій статті, ми коротко презентуємо нашу планету вірусів, після чого звернемося до великого та важливого питання в біології: чому більша частина життя є вірусною? Ключовий момент полягає в тому, що під час інфекційного циклу початковий віріон повністю розпадається і тільки асоційована з ним інформація передається наступному поколінню. У цьому полягає відмінність від клітинних організмів, котрі повинні передавати деяку фізичну частину самих себе від покоління до покоління. Спираючись на це припущення, пропонується визнати, що термодинамічні наслідки фізичної інформації (тобто, принцип Ландауера) спостерігаються у природних вірусних популяціях. Цей зв’язок між фізичною та генетичною інформацією, потім, використовується для побудови Вірусної Інформаційної Гіпотези, котра стверджує, що генетична інформація відтворює себе за рахунок енергетичної ефективності системи (тобто, є вірусною за своєю сутністю). Нарешті, ми показуємо яким чином вірусна інформація може бути перевірена та ілюструємо як цей новий погляд може пояснити існуючі екологічні й еволюційні теорії, спираючись на більш фундаментальні принципи.
Віруси є визначними рушіями глобальної біогеохімії та етіологічними агентами багатьох хвороб. Вони також є переможцями у грі життя: на планеті вірусів більше, ніж клітинних організмів, і вони представляють більшу частину генетичного розмаїття планети. У дійсності, є підстави вважати життя вірусним інкубатором. Однак, більшість екологічних та еволюційних теорій, що були розвинуті й продовжують розвиватися, не беруть до уваги віросферу. Це означає, що ці теорії потребують повторної інтерпретації у світлі знань про віруси, або ж ми маємо розробити нову теорію з вірусологічної точки зору. У цій статті, ми коротко презентуємо нашу планету вірусів, після чого звернемося до великого та важливого питання в біології: чому більша частина життя є вірусною? Ключовий момент полягає в тому, що під час інфекційного циклу початковий віріон повністю розпадається і тільки асоційована з ним інформація передається наступному поколінню. У цьому полягає відмінність від клітинних організмів, котрі повинні передавати деяку фізичну частину самих себе від покоління до покоління. Спираючись на це припущення, пропонується визнати, що термодинамічні наслідки фізичної інформації (тобто, принцип Ландауера) спостерігаються у природних вірусних популяціях. Цей зв’язок між фізичною та генетичною інформацією, потім, використовується для побудови Вірусної Інформаційної Гіпотези, котра стверджує, що генетична інформація відтворює себе за рахунок енергетичної ефективності системи (тобто, є вірусною за своєю сутністю). Нарешті, ми показуємо яким чином вірусна інформація може бути перевірена та ілюструємо як цей новий погляд може пояснити існуючі екологічні й еволюційні теорії, спираючись на більш фундаментальні принципи.
Віруси як інформація
Віруси – це єдині біологічні сутності, котрі відтворюються винятково як інформація. Коли вірус потрапляє в клітину організму хазяїна, віріон повністю дезінтегрується і нуклеїнова кислота копіюється для створення нових геномів, які потім пакуються та вивільняються у вигляді нових віріонів. Фізично, немає нічого, що передавалося б від початкового віріону наступним поколінням. Жодна молекула, атом чи кварк не повинні переноситися між старим на новим. Єдина річ, яка має переміщатися між вірусними поколіннями – це інформація, необхідна для побудови наступної генерації вірусів. Решта біологічних об’єктів оперує іншими механізмами. Кожна нова клітина фізично поділяє усі свої молекули з початковою материнською клітиною на момент поділу.
Тут ми пропонуємо Вірусну Інформаційну Гіпотезу, яка доводить, що:
- Фізична інформація стосується положення у Всесвіті.
- Біологія створює фізичну інформацію шляхом зміни положення (просторового розташування) матерії, ефективно працюючи за принципом Максвелівського демона.
- Вірусна інформація конвертує різні типи фізичної інформації у себе за рахунок загальної енергетичної ефективності.
- Існує термодинамічна ціна руйнування фізичної інформації, котра кількісно визначається за принципом Ландауера. Надзвичайно великі популяції, на кшталт вірусних, зазнають добору на межі Ландауера і це можна спостерігати.
Ласкаво просимо до світу вірусів
Люди спостерігають природу в середніх масштабах (тобто, від міліметрів до кілометрів). Наш мозок добре справляється з обробкою даних такого штибу, – від спостереження квітування черешневих дерев до пірнання біля коралових рифів, до вимірювання дзьобів зябликів. Але наші чуття зводять нас на манівці, адже донедавна ми недобачали більшу частину життя. На черешневому дереві квітки, коріння, гілки та листя мають мільйони вірусів та їхньої здобичі у вигляді бактерій. Пропливаючи над кораловим рифом, слід знати, що кожен мілілітр морської води є домівкою для десяти мільйонів вірусів і кожна поверхня, включаючи слиз на коралах та рибах, вкрита ще більшою кількістю вірусів. А значна частина ДНК, що літає в організмах славетних дарвінівських в’юрків [1], насправді належить бактеріям та вірусам.
Вірусів особливо легко недобачити, оскільки вони перебувають повністю за межею нашого сенсорного сприйняття. Це проблема, тому що випускаючи з поля зору віросферу біологи насправді ігнорували найбільш різноманітні та численні біологічні сутності на Землі. За дуже обережними оцінками, їх є 1х1031. Це базується на оцінці кількості мікроорганізмів на планеті приблизно у 1х1030 та на тому припущенні, що, в середньому, на одну прокаріотичну клітину випадає 10 вірусів. Якби інопланетянин відвідав нашу планету й мав інший рівень сенсорного сприйняття, який дозволяв би йому безпосередньо виявляти віруси, то він, швидше за все, вважав би їх домінантною формою життя. (Примітка для читача: якщо ви обізнані у галузях історії та біології вірусів, то можете спокійно проминути наступну частину розділу, оскільки у ній ми обговорюватимемо ці питання).
Звідки ж ми знаємо про існування такої великої кількості вірусів? Спочатку їх рахували використовуючи електронний мікроскоп. Зараз же їх зазвичай рахують використовуючи епіфлуоресцентну мікроскопію. Наприклад, для підрахунку кількості вірусів, що містяться в одному мілілітрі морської води, зразок пропускається крізь скляний фільтр з порами, діаметр яких становить 0,02 мкм (вони достатньо маленькі для уловлення вірусів). Потім фільтр обробляється мітками для ДНК, світіння яких можна помітити за допомогою флуоресцентного мікроскопа. Технічно, тим, що біологи рахують у дійсності, є вірусоподібні часточки (ВПЧ). ВПЧ – це щось, що виглядає як вірус, але не було формально охарактеризоване у такій якості і не було показано, що воно діє як вірус; тобто, інфікує і потім реплікується у клітині хазяїна. Навіть беручи до уваги неймовірно малі розміри вірусів, 1031 – це щось величезне. Якщо б ви розташували усі віріони в одну лінію, то її довжина була б достатньою для того, щоб тисячу разів обмотати нашу галактику.
Якщо загальна кількість вірусів є надзвичайно великою, то дійсно неймовірною є їхня динаміка. За нашими найкращими підрахунками, щотижня 1031 вірусів гине і продукується 1031 нових їм на заміну. Це означає, що приблизно 1,7х1025 нових вірусів утворюється щосекунди. Для кожного нового вірусу, приблизно, слід синтезувати 50 000 пар нуклеотидів ДНК. Таким чином, щосекунди більше ніж 1030 пар нуклеотидів вірусної ДНК синтезується на планеті Земля. Оскільки переважна більшість цих вірусів уражує мікробів (бактерії та археї, два з трьох доменів життя), поява цих вірусів означає загибель приблизно 1024 мікробних клітин щосекунди. Це посилює розмаїття мікроорганізмів та продуктивність екосистем. Це також вагомий фактор у глобальній циркуляції енергії та поживних речовин. Мета цих викладок полягає у тому, щоби показати наскільки численними, ємнісними та динамічними насправді є ці 1031 вірусів. Коли йдеться про віросферу, надзвичайно малореалістичні події стають ймовірнісними певностями.
Якщо навіть віруси і є домінантами у нашому закутку Всесвіту, більшість людей зважає на них лише тоді, коли вони спричиняють якісь хвороби [2]. Але насправді, більшість вірусів є фагами: тобто, вірусами, які уражують бактерій. В 1915 році англієць Фредерік Туорт відкрив “ультрамікроскопічний вірус”, що перетворює бактерії на красиві гранули. У його використанні, слово “вірус”, як видається, означало просто “інфекційний агент”. Він писав про “дуже дрібну бактерію, яка житиме тільки зростаючи на живому матеріалі… чи це форма життя нижчого рівня організації, ніж бактеріальний”. Вірус був знищений при 600С, його можна було культивувати тільки на бактеріях. “В цілому, видається ймовірним, хоча й немає певності, що активний прозорий матеріал продукується мікрококами, а потім призводить до їхньої власної загибелі і може передаватися здоровим свіжим культурам, його можна розглядати як збудника гострої інфекційної хвороби мікрококів”. Ось так бактерія стає хворою.
Канадець французького походження, Фелікс д’Ерель, пішов далі і показав, що здатний до фільтрації “антагоністичний мікроб”, який може вбивати бактерію Shygella dysenteriae, виділявся з пацієнтів, котрі хворіли на ентерит, що розвивався після захворювання на дизентерію. Він здійснив перший аналіз бляшок і показав, що титри цього агенту були найвищими під час одужання пацієнта. Культивування цього агента потребувало наявності живих дизентерійних бактерій, але за таких умов агент міг культивуватися протягом 50 послідовних пасажів. Д’Ерель писав: “зникнення дизентерійної бацили співпадає з появою невидимого мікроба… Цей мікроб, справді мікроб імунності, є облігатним бактеріофагом” – перше використання терміну.
Бактеріофаг означає “пожирач бактерій”, як правило, у вжитку термін скорочується і говорять просто “фаг”. Це підклас вірусів, що уражують бактеріальний домен життя. На зорі вірусології, мисливці за вірусами швидко збагнули, що фаги дуже розмаїті, кожен з них є вибагливим, щодо того, якого хазяїна уражувати. Така специфічність фагів до специфічних штамів бактерій була одним з ранніх шляхів мікробіологічної ідентифікації, яка здійснювалася через процедуру, котра називалася визначення типу фагів. Принципово, це може бути зроблено шляхом культивування бактерій у тестовій пробірці й подальшого додавання різних бактеріофагів. Якщо вміст пробірок стає прозорим, це означає, що всі бактерії були вбиті бактеріофагом. Використовуючи цей підхід, були класифіковані тисячі бактеріофагів та штамів їхніх хазяїв. Визначення таксономічної належності хазяїна є одним з найбільш плідних підходів для опису віросфери.
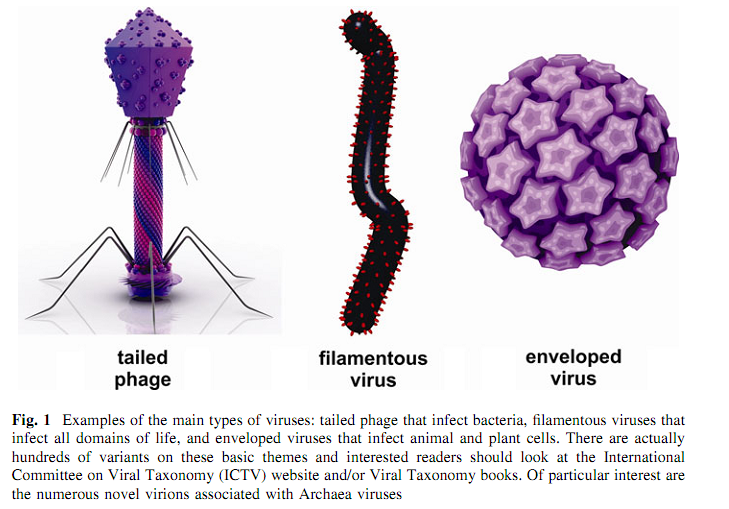
Інший шлях опису вірусів полягає у тому, щоб їх побачити за допомогою електронного мікроскопа. Віруси поза клітинами організму хазяїна називаються віріонами і вони є одними з найдивовижніших створінь, які коли-небудь були відкриті. Типовий бактеріофаг виглядає неначе якийсь місяцехід, він має білковий капсид, що захищає геном у вигляді подвійного ланцюжка ДНК. Він має хвіст, котрий використовується для впорскування вірусного геному в клітину хазяїна, а фібри хвоста допомагають бактеріофагу знайти правильного хазяїна. Вірусний капсид, зазвичай, має один з двох базових типів структури: паличковий чи ікосаедричний, розмір же його може значно варіювати, а геном, який пакується у нього, також може мати різний розмір, відрізняючись у різних вірусів більше ніж у 30 разів. При формуванні паличкоподібних часточок, білки капсиду вкладаються по спіралі навколо вірусної ДНК чи РНК. Вірус тютюнової мозаїки (ВТМ) є класичним прикладом цього типу симетрії. Іншим розповсюдженим типом структури капсиду є ікосаедр, що оточує серцевину з нуклеїнової кислоти. Серед понад п’яти тисяч бактеріофагів, котрі були описані і розглянуті за допомогою електронного мікроскопа станом на 2000 рік, 96% з них є “бактеріофагами з хвостом”, що складаються з ікосаедричної головки, яка містить геном, та мають хвіст, що функціонує для ідентифікації хазяїна і доставки генома всередину клітини. Хвостові структури поділяють схожих на місяцеходи бактеріофагів на три головні групи: λ-подібних фагів, котрі мають довгий, гнучкий хвіст; Т7-подібних фагів, які мають короткий та скоротливий хвіст; і Т4-подібних фагів, що мають довгий скоротливий хвіст (Аккерман, 2007). Часто капсидні білки та нуклеїнові кислоти геному збираються самі in vitro та формують інфекційні віріони. Не треба ні додаткової інформації, ані джерела енергії. Віруси, котрі уражують тварин та рослини, часто мають вигляд ікосаедричної структури, оточеної ліпідним суперкапсидом.
Віріони – це вишукано організовані хижаки, котрі знаходять та вбивають своїх хазяїв. Загалом, віріони мають невеликий негативний заряд, тож вони відштовхуються один від одного під час лізису клітини-хазяїна. Це дозволяє їм поширюватися та уникати скупчення у вивільненому вмісті клітини, яка зазнала лізису. Якщо говорити точніше, то видається, що заряди розташовані таким чином, що віріони насправді є диполями; тобто, мають негативний заряд навколо голівки капсиду і незначний позитивний заряд на хвостах. Це, як припускається, орієнтує їх хвостами вперед під час нападу на бактеріальну клітину-хазяїна (яка має невеликий негативний заряд).
Під час фази атаки, віріон, перш за все, електростатично притягується до поверхні клітини. Він котиться зовні по ній, а хвіст шукає специфічні рецептори. Якщо ця клітина є потрібним бактеріальним хазяїном, то бактеріофаг знайде рецептор та приєднається до нього. Коли це трапляється, хвіст бактеріофага просвердлить мембрану клітини і клітинну стінку, й вірусний геном отримає змогу потрапити в цитоплазму. Для досягнення цього, зовнішня частина скоротливого хвоста перебудовує свою молекулярну структуру таким чином, щоб рурка всередині оболонки могла проколоти клітину. Це дозволяє ДНК впорскуватися з неймовірною силою. Цей процес не так вже й відрізняється від вторинних щелеп у фільмі Рідлі Скота “Чужий”; він жахливий, якщо б вам довелося бути розміром з мікроба.
На додачу до хвостів, капсиди бактеріофагів часто декоровані вторинними структурами, що полегшують атаку. До них належать гачки, котрі загарбують бактеріальні джгутики, щоб фаг, таким чином, притягувався до бактерії-хазяїна. Інші молекулярні аксесуари, ймовірно, допомагають віріону пережити різні умови довкілля чи діють як камуфляж, що дозволяє уникнути захисних ферментів, вироблюваних хазяїном. Постійна війна між здатностями віріонів знаходити й інфікувати клітини та силами протидії хазяїна зумовлює еволюційну динаміку, відому під назвою Гіпотези Червоної Королеви (Ван Вален, 1974) та екологічні цикли, котрі називаються моделлю Лотка-Вольтерра (“Вбий переможця”).
Існує декілька проблем, які необхідно вирішити для вивчення розмаїття та динаміки глобального вірому. Для культивування вірусу ви маєте вирощувати його хазяїна, а на сьогодні ми маємо розроблені методики для культивування приблизно 1% мікроорганізмів довкілля. А якщо умови для культивування мікроорганізму встановлені, їх слід модифікувати для того, щоб стимулювати вірусну інфекцію. Зважаючи на ці виклики, шлях культивування буде складним та непривабливим. А як щодо секвенування вірусної ДНК? Секвенування гену 16S рибосомної РНК (рДНК) є поширеною технікою, що використовується для аналізування різноманітності популяцій мікроорганізмів, і вона користується високою консервативністю цього одного гена серед майже всіх мікробів, таким чином, вдається уникнути порівняння цілих геномів для вивчення розмаїття популяцій. Однак, неможливо застосувати такий же підхід щодо вірусів, оскільки у них немає такого гена, який був би спільним для всіх груп. Для того, щоб обійти це обмеження, була розроблена методика вибіркового секвенування рідкісних фрагментів з пулу усіх вірусних геномів популяції. Цей підхід називається метагеномікою.
Аналізування усього генетичного пулу (метагеному) зразка було уперше виконане на вірусних спільнотах, виділених з морської води в Сан-Дієго (Брейтбарт та ін., 2002). Це раннє дослідження показало, що переважна більшість вірусних послідовностей (80% чи навіть більше) не пізнавалися при використанні звичних методів біоінформатичного пошуку. Іншими словами, вірусні ДНК вірусів, що не культивувалися, були настільки відмінними від будь-якої відомої послідовності вірусного, бактеріального чи еукаріотичного походження, – інформація про які накопичена у різних базах даних (наприклад, GenBank), – що у нас не було жодних ідей стосовно того, що вони роблять чи кому вони належать. Не дивлячись на те, що неймовірний обсяг даних про секвеновані послідовності додався до баз даних з того часу, коли були здійснені перші дослідження у галузі метагеноміки, більшість вірусів залишається невідомою. З іншого боку, метагеноміка мікроорганізмів, яка одразу ж зародилася після метагеноміки вірусів, є куди менш дивовижною – лише близько 20% секвенованих послідовностей не мають жодних відповідностей у базах даних (Дінсдейл та ін., 2008). Оскільки вірусів дуже багато, набагато більше ніж мікроорганізмів, і оскільки більшість інформації, що міститься у вірусних геномах є невідомою, віруси є фінальним прикордонням недослідженої генетичної різноманітності і є найбільшим з існуючих генетичним вмістилищем. У нас залишається запитання: чому існує так багато вірусів?
Демони та інформація
Досі ми стверджували, що віруси надзвичайно численні, неймовірно різноманітні і що вони подорожують крізь час і простір як інформація. Ми постулюємо, що цей зв’язок між вірусами та інформацією є ключовим для їхнього успіху, але що означає поняття “інформація”? У сенсі спілкування, інформація є мірою “здивування”. Чим більшим є подив при спостереженні об’єкта, тим більше інформації він містить. Золото містить більше інформації, ніж водень (тобто, знаходження золота викликає більший подив, його важче знайти). Розглядаючи цю концепцію ретельніше, стає зрозумілим, що інформація, насправді, це визначення позиції (просторового розташування) у Всесвіті. Таким чином, золото створене шляхом стискання протонів, нейтронів та електронів разом у просторі й часі. А коли ці часточки замикаються разом, ступені свободи втрачаються і створюється унікальна, а тому високоінформативна, подія. Така організація матерії у часі й просторі є фізичною інформацією.
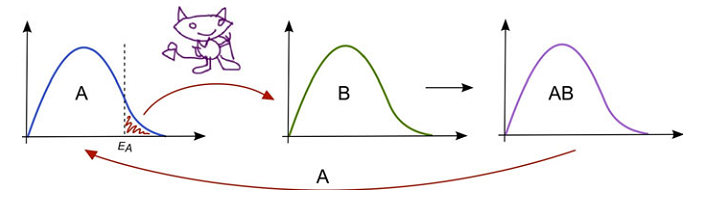
Рис. 2. Відображення демона Максвелла та принципу Ландауера. Демон/фермент вибірково підбирає молекули “А”, котрі мають достатню енергію для взаємодії з реагентом “В”, що призводить до утворення продукту “АВ”. Цей процес призводить до незначного охолодження сукупності “А”. Ця втрата теплоти компенсується системі за рахунок оточуючого Всесвіту. Під час деградації/розмивання “АВ”, “А” повертається назад до своєї сукупності і ця теплота може бути виміряна при застосуванні таких методів, як ізотермічна калориметрія.
Фізична інформація не створюється задарма. Термодинамічний наслідок фізичної інформації уперше був математично визначений професором Рольфом Ландауером, який підрахував, що мінімум енергії (Е), котра зберігається в одному біті інформації, дорівнює kTln, де k – стала Больцмана, а Т – температура за шкалою Кельвіна (Ландауер, 1996). Теплота, котра вивільняється внаслідок знищення фізичної інформації, найкраще може бути уявлена, якщо звернутися до демона Максвелла. Спочатку презентований у якості виклику для другого закону термодинаміки, демон є гіпотетичним створінням, яке може підбирати “гарячі” молекули з одного контейнера та переміщувати їх до іншого. Це створює температурну різницю, яка може бути використаною для роботи якогось двигуна. Отже, маючи такого демона, ми можемо створити такий собі вічний двигун. Саме Лео Сцілард показав причину того, чому цього не трапляється у дійсності: адже демон, насправді, отримує інформацію про відносне просторове розташування молекул. Таке усвідомлення вбило вічний двигун і зліквідувало виклик Максвелла другому законові термодинаміки.
Тепер уявімо демона Максвелла біохімічною системою (рис. 2). За певної температури, реагуючі молекули “А” мають різні швидкості, як це описується Больцманівським розподілом. Найшвидші/найгарячіші молекули “А” знаходяться праворуч у цьому розподілі. Для наших цілей, молекули, енергія яких є вищою від енергії активації (ЕА), мають достатню швидкість для участі в хімічній реакції. Тепер уявімо демона Максвелла, який вибірково підхоплює молекули “А” із сукупності ЕА та переміщує їх до другого реагуючого пулу “В”. Це створює продукт та ефективно вловлює обидві молекули в продукті “АВ”. Роблячи таке, демон збільшує інформацію системи. Коли “АВ” розпадається на складові компоненти, “А” надійде до своєї початкової сукупності та підігріє її [3]. Це збільшення температури описується принципом Ландауера.
Ми постулюємо, що біологія поводиться як демон Максвелла, коли у ролі демонів виступають ферменти, які вибірково підхоплюють молекули ЕА для утворення продуктів. Це створює фізичну інформацію, яка може бути використаною для здійснення роботи. Одне застереження стосовно цієї схеми робота – завдяки – інформації полягає в тому, що вона потребує риштування на кшталт комп’ютера. Ми припускаємо, що генетична інформація є набором інструкцій для побудови риштування для демонів Максвелла, котрі перетворюють різні типи фізичної інформації на збільшення чисельності самих себе. Ця нова інформація має термодинамічну ціну, коли вона руйнується і певна кількість теплоти виділяється внаслідок знищення інформації – це також описано принципом Ландауера. Повинно бути можливим спостереження зв’язку між фізичною інформацією та термодинамікою і використання останньої для кращого розуміння біології в цілому, та успіху вірусів зокрема.
Вірусна інформація проти фізичної інформації
Давайте порівняємо та протиставимо фізичну й вірусну інформацію. Сила тяжіння організовує фізичні властивості Всесвіту. Тяжіння згромаджує матерію, яка збільшує важливість трьох інших фундаментальних взаємодій. Організовуючи матерію у часі й просторі, гравітація створює фізичну інформацію. Хмара субатомних часточок з часів Великого Вибуху розширилася рівномірно по всьому Всесвіту. Натомість, маленькі недосконалості дозволили гравітації притягнути деякі часточки докупи; а вони притягнули інші. Сформувалися аккреційні диски, на їх основі утворилися зірки, де гравітація стягнула матерію докупи формуючи важчі елементи і призвела до появи електромагнітного випромінювання. Ці процеси збільшують ємність фізичної інформації у Всесвіті шляхом строгих фізичних явищ. Гравітація також посилює сама себе – більші речі притягують більше об’єктів, створюючи позитивний зворотній зв’язок. Біологія також сама себе посилює шляхом організації компонентів та їхньої концентрації. Точно як гравітація, життя створює організацію часточок у Всесвіті шляхом накладання та перестановок. Організація матерії біологією призводить до вірусної інформації, тому що вона перетворює фізичну інформацію на себе за рахунок максимальної ефективності з термодинамічної точки погляду.
Вірусна інформація й решта біології
Два первинних джерела фізичної інформації використовуються для перетворення на вірусну інформацію: електромагнітне випромінювання внаслідок злиття (підвалина фотосинтезу) та окиснення, що призводить до появи побічних продуктів внаслідок розщеплення (підвалина хемотрофії). Давайте перевіримо найбільший біом на землі (потенційно), глибоку й гарячу біосферу, що існує в межах земної кори, у якості прикладу того, яким чином може працювати зворотній зв’язок вірусної інформації. У цій екосистемі є дуже просте джерело енергії, – розщеплення води -, яке дає енергію для продукування клітинної біомаси. З урахуванням температури й тиску у цій системі, єдиними відомими хижаками серед мікробних її мешканців є віруси. У цій простій харчовій мережі, головні “правила” екології та еволюції є очевидними. Це (1) виключення ніші, (2) створення ніші та (3) динаміка Червоної Королеви/Вбий-Переможця, яка дуже яскраво виявляється та зумовлюється (4) вірусною інформацією.
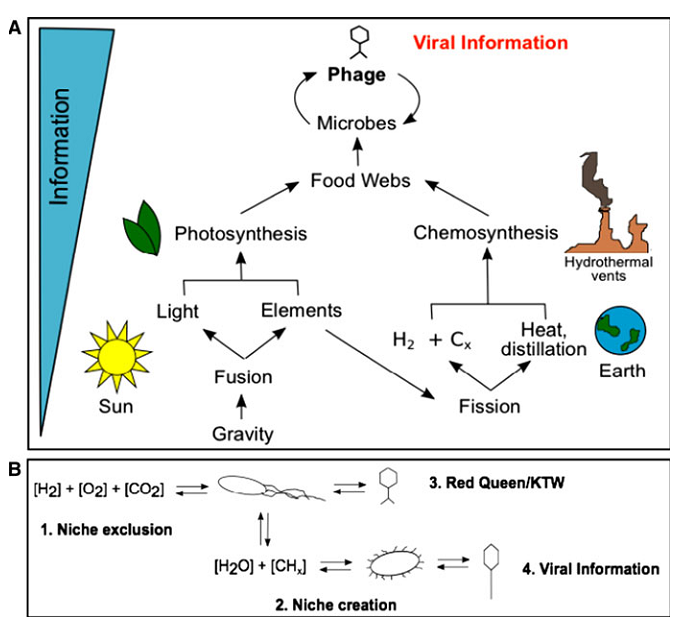
Рис. 3. Від гравітації до вірусної інформації: від пилу до фагів. а Схематичне зображення того, як гравітація призводить до появи вірусної інформації. b Схематичне відображення того, як віруси формують екологію (1 – 3) та еволюцію (3), призводячи до диверсифікації та збільшення вірусної інформації (4).
На першому етапі, популяція мікроорганізмів живе за рахунок ресурсів місцевого довкілля (у даному випадку, розщеплення води). Це призводить до виснаження цього ресурсу в системі, виникнення конкуренції та виключення ніші; мікроорганізми, які використовують ці ресурси найшвидше, перемагають. У той же час, віруси в системі суттєво карають найуспішніших мікроорганізмів вбиваючи їх. Спричинюваний вірусами лізис призводить до вивільнення вмісту клітин у навколишнє середовище, і тоді починають процвітати нові мікроорганізми, які використовують цей новий ресурс, вони витісняють інших з цієї нової ніші, створюючи новий набір решток та ресурсів і підтримуючи нову популяцію вірусів. Фактично, віруси самі створюють умови для власного відтворення.
Тиск хижаків також призводить до того, що мікроорганізми частково змінюються, щоб уникнути впізнання вірусами (тобто, має місце динаміка Червоної Королеви, або бігу для того, щоб залишатися на місці), тоді як віруси адаптуються для впізнання нових мікроорганізмів.
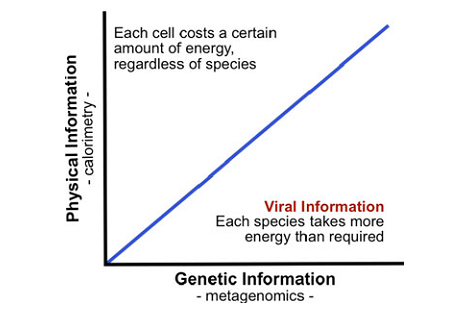
Рис. 4. У пошуках вірусної інформації. Лінія відображає де саме кількість фізичної та генетичної інформації, що міститься в клітинах, є однаковою. Спільноти над лінією містять більше фізичної інформації, ніж генетичної, через низьку генетичну різноманітність (мало видів, зате багато особин), а кожна особина вимагає певної кількості енергії, незважаючи на свій генетичний склад. Спільноти під лінією містять більше генетичної інформації. Саме тут енергетична ціна інформації стає очевидною, і тут ми сподіваємося знайти вірусну інформацію.
Іншими словами, віруси рухають еволюцію геномів мікроорганізмів та ніш, призводячи до виняткового збільшення вірусної інформації. Оскільки віруси є більш розмаїтими, на них випадає створення найбільшої кількості нової генетичної інформації, яка потім може передаватися хазяїнам-мікроорганізмам шляхом горизонтального перенесення генів (ГПГ). У кінцевому рахунку, спільнота перетворює фізичну інформацію на генетичну. Ми припускаємо, що цей етап є вірусним тому що він здійснюється за великої термодинамічної неефективності; тобто, продукується велика кількість теплоти, яка втрачається. Використовуючи правило піраміди, згідно з яким кожне трофічне перенесення супроводжується втратою 90% енергії, кожен джоуль вірусної інформації коштує системі 100 джоулів інформації фізичної.
Вимірювання вірусної інформації
Деструкція фізичної інформації, як обговорювалося вище, зумовлює вивільнення теплоти, згідно з принципом Ландауера. Ця теплота може бути виміряна шляхом калориметрії. Конкретно, ізотермічна калориметрія говорить нам про рівень перетворення фізичної інформації сукупності на теплоту [4]. Генетична інформація цієї ж сукупності (спільноти) також може бути виміряною, у цьому випадку – шляхом секвенування ДНК. Спираючись на ці дві методики, ми пропонуємо наступний експеримент, під час якого фізична інформація відстежується за допомогою калориметрії, а генетична – метагеноміки. Коли обидва методи суміщаються, як показано на рис. 4, ми постулюємо, що сукупність, у якій домінує вірусна інформація, опиняється у правому нижньому секторі графіка, де генетична інформація створюється коштом термодинамічної ефективності (тобто, низького рівня перетворення на фізичну інформацію).
Чи є якісь свідчення того, що вірусна інформація реальна? Джамалі з колегами використали ізотермічну калориметрію для вивчення вивільнення теплоти морськими мікроорганізмами та вірусними спільнотами (Джамалі та ін., 2012). У цьому експерименті, віруси знизили питому частку клітинного компоненту приблизно на 25%. У той же час, віруси збільшили вихід роботи системи на 200%. Зменшення кількості клітин, у поєднанні зі збільшенням різноманітності, виглядає дуже схожим на вірусну інформацію. Майбутні експерименти такого штибу запропонують критерії для перевірки Вірусної Інформаційної Гіпотези.
Спостереження вірусної інформації в природі
У якості одного з можливих наслідків вірусної інформації в природі, давайте розглянемо глобальне збереження вірусних послідовностей (сиквенсів). Специфічні ПЛР “полюють” на однакових вірусів та/чи вірусні гени, котрі показують, що ці віруси/вірусні гени є відносно поширеними у всьому світі. Наприклад, ПЛР праймери були сконструйовані таким чином, щоб специфічно ампліфікувати дві вірусні послідовності, названі HECTOR та PARIS. Ці, так звані PUP-послідовності (полімерази з некультивованих подофагів) були представлені у більшості середовищ, що досліджувалися; була встановлена їхня суттєва ідентичність (99% на нуклеотидному рівні). Подібно, метагеномні зразки були визнані цілком ідентичними, – вірусні послідовності перекривалися у зразках, узятих у різних віддалених частинах океану. Нарешті, вивчення геному бактеріофагів виявило значні відповідності нуклеотидних послідовностей у дуже різних геномах фагів (Грехем Хатфул, особисте спілкування).
Значна поширеність майже ідентичних послідовностей (нуклеотидних) по всій планеті потребує пояснення. Ми припускаємо, що така надзвичайно послідовна глобальна консервація зумовлена енергетичною вартістю знищення інформації. Як ми бачили, на планеті існує майже астрономічна кількість вірусів. Оцінюється, що кожна вірусна популяція (тобто, кількість представників одного виду) становить 1023. Якщо кожен вірус у популяції відрізнявся від інших лишень одним бітом інформації, тоді теплота, яка вивільнилася б внаслідок знищення цієї додаткової інформації, становила б 1800 джоулів згідно з принципом Ландауера (рис. 5). Іншими словами, вірусна популяція, яка має одну мутацію на геном, потребує на 1,8 кДЖ більше енергії для реплікації кожного геному, ніж та популяція, де такої мутації немає. Протягом року, кількість енергії, яку потребує мутантна вірусна популяція у порівнянні з немутантною, становить близько 100 кДЖ, виходячи з припущення, що уся популяція оновлюється щотижня. За мільярд років, це становить 1014 Дж, що, при грубому порівнянні, еквівалентно кількості енергії, яка вивільняється під час вибуху атомної бомби [5]. Енергетично ефективні популяції перемагають тих, які є менш ефективними; тож, популяції вірусів зі зменшеними частотами мутацій перемагатимуть тих, котрі мають вищі частоти, за однаковості усіх інших чинників.
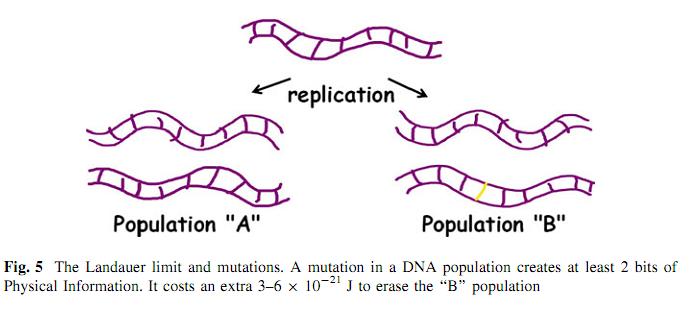
Додаткова енергетична вартість фізичної інформації, асоційованої з мутацією, може пояснити, чому ідентичні вірусні послідовності спостерігаються у глобальному масштабі. Фізична інформація у сенсі мутації є надзвичайно малим селективним тиском і ми припускаємо, що ліміт Ландауера є найменшою силою добору. Завдяки їхньому “суто інформаційному” стилю життя, легше спостерігати термодинамічні наслідки інформації у вірусних спільнотах [6]. Понад те, ми спостерігаємо це тільки в Природі, тому що надзвичайно важко виростити 1023 бактеріофагів (чи будь-яких інших біологічних об’єктів) у лабораторних умовах.
Висновок
Сприйняття біосфери як масивної системи, яка, в кінцевому рахунку, підтримує (“живить”) віруси, допомагає нам знайти підхід для вирішення великого й видатного питання: чому віруси є домінантами біологічного розмаїття? Це питання могло навіть не поставати перед біологами більш ранніх часів, просто через те, що вони не знали справжніх розмірів віросфери. Сучасна біологія, проте, потребує інкорпорації цього природного феномену у свій канон. Вірусна Інформаційна Гіпотеза має потенціал для синтезу екології й еволюційної теорії шляхом поєднання вірусів з рештою біології у рамках термодинаміки.
Форест Рохвер,
Кеті Беротт
(Університет Сан-Дієго)
Джерело: Springer
Переклад з англійської: Юрій Косенко
P.S. Список літературних джерел та іншу супутню інформацію можна побачити перейшовши за вищевказаним посиланням.
[1] Зробивши деякі чорнові розрахунки для зебрового в’юрка Taeniopygia guttata, ми оцінили, що близько 10% усієї ДНК в організмі пташки належить вірусам та бактеріям.
[2] Намагання ідентифікувати бактерію – збудника хвороби тютюну – призвело до виявлення першого вірусу Бейєрінком у 1898 році. Інфекційний агент був описаний, як “заразний живий флюїд”, що фільтрується, пізніше його назвали вірусом тютюнової мозаїки. (На Заході пріоритет Д. Івановського, загалом, не визнається, – Прим. пер.).
[3] Іноді про таке легше думати, уявляючи собі тільки дві молекули А. Коли одна молекула забирається, система стає холоднішою. А коли ця молекула А повертається до системи, вона стає гарячішою.
[4] У цьому сенсі, фізична інформація становить 1\S, де S – ентропія. Ми віддаємо перевагу використанню терміна “інформація” у даному контексті, оскільки він краще пояснює концепцію. Однак, цілком прийнятно виражати її використовуючи поняття ентропії, S.
[5] Слід пам’ятати, що таке змагання відбувається, у дійсності, локально і серед малих часток усієї популяції.
[6] Віруси можуть бути не єдиною сферою, де можна спостерігати ліміт Ландауера. Ультраконсервативні елементи (УКЕ) трапляються у рослинах та тваринах, – у їхніх геномах. Існує 481 УКЕ (200 п.о.), спільних для людини, миші та щура, – це свідчення 300 мільйонів років еволюції. Ренекер та його команда ідентифікували велику кількість УКЕ (понад 100 п.о.), що є спільними для рослин і тварин (Ренекер та ін., 2012).